煙酰胺單核苷酸(NMN)可增加糖尿病前期女性的肌肉胰島素敏感性 (2021/01)
在囓齒動物中,肥胖和衰老會損害煙酰胺腺嘌呤二核苷酸 (NAD+) 的生物合成,從而導致代謝功能障礙 (1-8)。煙酰胺單核苷酸 (NMN) 可用性是哺乳動物 NAD+ 生物合成的限速因素。我們進行了一項為期 10 週的隨機、安慰劑對照、雙盲試驗,以評估補充 NMN 對超重或肥胖的絕經後前驅糖尿病婦女代謝功能的影響。使用高胰島素-正常血糖鉗評估胰島素刺激的葡萄糖處理,補充 NMN 後骨骼肌胰島素信號(AKT 和 mTOR 的磷酸化)增加,但安慰劑治療後沒有變化。 NMN 補充劑上調了血小板衍生生長因子受體 β 和其他與肌肉重塑相關的基因的表達。這些結果表明,NMN 增加了超重或肥胖的前驅糖尿病女性的肌肉胰島素敏感性、胰島素信號傳導和重塑。 (ClinicalTrial.gov NCT03151239)
一句話總結:煙酰胺單核苷酸(NMN)增強胰島素作用
煙酰胺腺嘌呤二核苷酸 (NAD+) 是 NAD+ 消耗酶的共底物,在調節多種生物過程中必不可少。在囓齒動物中,肝臟、脂肪組織和骨骼肌中 NAD+ 生物合成不足會導致肥胖和衰老相關代謝異常的發病機制,包括胰島素抵抗、β 細胞功能障礙和肝脂肪變性 (1-9)。 NAD+ 生物合成的主要途徑包括將煙酰胺轉化為煙酰胺單核苷酸 (NMN) 以及隨後將 NMN 轉化為 NAD+(5)。 NMN的產生是哺乳動物NAD+生物合成的關鍵限速因素。在餵食高脂肪飲食的肥胖小鼠中全身性 NMN 給藥會增加組織 NAD+ 濃度並改善葡萄糖耐量、胰島素敏感性和 β 細胞功能 (1, 4, 10),並且在定期餵食的小鼠中長期 NMN 給藥可以減輕年齡-相關的胰島素抵抗 (11)。 NMN補充劑對囓齒動物的有益作用導致NMN產品的快速商業化開發,NMN作為改善葡萄糖控制、增強能量代謝和逆轉衰老代謝並發症的補充劑在美國和其他國家上市。儘管 NMN 存在於天然食品中,包括毛豆、西蘭花、鱷梨、西紅柿和牛奶,但作為健康飲食的一部分,通常每天消耗的 NMN 量可能少於 2 毫克/天 (5)。商業產品中 NMN 的劑量範圍為 50 毫克至 150 毫克/膠囊,一些消費者每天服用兩粒 150 毫克膠囊。然而,我們不知道關於每日補充 NMN 對人體代謝影響的研究。
我們對超重或肥胖(體重指數 25.3 至 39.1 kg/m2)的絕經後婦女進行了為期 10 週的隨機、安慰劑對照、雙盲試驗,以確定補充 NMN 對以下方面的影響:組成,ii) 骨骼肌胰島素敏感性和胰島素信號傳導; iii) 肌肉 NAD+ 含量和整體基因表達譜。我們專門研究了絕經後婦女,因為 NMN 治療對患有飲食誘導糖尿病的雄性小鼠產生的有益代謝影響大於雄性小鼠 (1)。
根據美國糖尿病協會 (12) 提出的標準,25 名超重或肥胖的絕經後糖尿病前期婦女完成了這項研究; 12 人被隨機分配到安慰劑組,13 人被隨機分配到 NMN 組(250 mg/天;Oriental Yeast Co., Ltd.,Tokyo,Japan)(圖 S1)。基線測試包括: i) 雙能 X 射線吸收測定法和磁共振成像以評估身體成分; ii) 高胰島素-正常血糖鉗夾程序(4 小時基礎階段和 4 小時 50 mU/m2 /min 的胰島素輸注),結合穩定同位素葡萄糖和棕櫚酸鹽示踪劑輸注,以評估多器官胰島素敏感性; iii) 基礎期股四頭肌經皮活檢和胰島素輸注,以評估 NMN 治療的細胞效應。基線研究完成後,參與者被隨機分配到安慰劑或 NMN 治療 10 週。在參與者完成約 10 週的治療後,重複在基線進行的所有研究程序。此外,在安慰劑或 NMN 攝入後 4 小時評估外周血單個核細胞 (PBMC) NAD+ 含量。根據每週的藥片計數,安慰劑組 99.6% 的處方藥和 NMN 組 100% 的處方藥被參與者服用。兩組均未報告不良事件,也未檢測到標準血液檢查異常(表 S1)。
N-甲基-2-吡啶酮-5-甲酰胺和N-甲基-4-吡啶酮-5-甲酰胺的血漿濃度是NMN產生的代謝物(13),在NMN治療10週後增加,但安慰劑沒有增加(圖1) . 1A)。 NMN 治療 10 週後,PBMC 中的基礎 NAD+ 含量增加,但安慰劑治療後沒有變化(圖 1B)。在治療 10 週結束時攝入單次 250 mg 劑量的 NMN 不會導致 PBMC NAD+ 含量進一步增加至高於基礎水平,這是在攝入後 240 分鐘獲得的連續血液樣本中評估的(圖 1C);然而,由於 NMN 組的基礎值較高,NMN 組的 240 分鐘 PBMC NAD+ 含量高於零的曲線下面積比安慰劑組高 43%(p<0.01)(圖 1D) )。在最後一劑安慰劑或 NMN 後 1.5 小時獲取股四頭肌組織樣本,以評估治療對骨骼肌 NMN 代謝物的影響。儘管兩組的肌肉 NAD+ 和煙酰胺含量在治療 10 週後都沒有變化(圖 1、 、EE 和 和 F)、F),但肌肉 N-甲基-煙酰胺、甲基-2-吡啶酮-5-甲酰胺、和 N-methyl-4-pyridone-5-carboxamide 在 NMN 而不是安慰劑治療後增加(圖 1, ,G,G, ,H,H 和 I),I),表明 NMN 治療增加了肌肉 NAD+周轉。
一句話總結:煙酰胺單核苷酸(NMN)增強胰島素作用
煙酰胺腺嘌呤二核苷酸 (NAD+) 是 NAD+ 消耗酶的共底物,在調節多種生物過程中必不可少。在囓齒動物中,肝臟、脂肪組織和骨骼肌中 NAD+ 生物合成不足會導致肥胖和衰老相關代謝異常的發病機制,包括胰島素抵抗、β 細胞功能障礙和肝脂肪變性 (1-9)。 NAD+ 生物合成的主要途徑包括將煙酰胺轉化為煙酰胺單核苷酸 (NMN) 以及隨後將 NMN 轉化為 NAD+(5)。 NMN的產生是哺乳動物NAD+生物合成的關鍵限速因素。在餵食高脂肪飲食的肥胖小鼠中全身性 NMN 給藥會增加組織 NAD+ 濃度並改善葡萄糖耐量、胰島素敏感性和 β 細胞功能 (1, 4, 10),並且在定期餵食的小鼠中長期 NMN 給藥可以減輕年齡-相關的胰島素抵抗 (11)。 NMN補充劑對囓齒動物的有益作用導致NMN產品的快速商業化開發,NMN作為改善葡萄糖控制、增強能量代謝和逆轉衰老代謝並發症的補充劑在美國和其他國家上市。儘管 NMN 存在於天然食品中,包括毛豆、西蘭花、鱷梨、西紅柿和牛奶,但作為健康飲食的一部分,通常每天消耗的 NMN 量可能少於 2 毫克/天 (5)。商業產品中 NMN 的劑量範圍為 50 毫克至 150 毫克/膠囊,一些消費者每天服用兩粒 150 毫克膠囊。然而,我們不知道關於每日補充 NMN 對人體代謝影響的研究。
我們對超重或肥胖(體重指數 25.3 至 39.1 kg/m2)的絕經後婦女進行了為期 10 週的隨機、安慰劑對照、雙盲試驗,以確定補充 NMN 對以下方面的影響:組成,ii) 骨骼肌胰島素敏感性和胰島素信號傳導; iii) 肌肉 NAD+ 含量和整體基因表達譜。我們專門研究了絕經後婦女,因為 NMN 治療對患有飲食誘導糖尿病的雄性小鼠產生的有益代謝影響大於雄性小鼠 (1)。
根據美國糖尿病協會 (12) 提出的標準,25 名超重或肥胖的絕經後糖尿病前期婦女完成了這項研究; 12 人被隨機分配到安慰劑組,13 人被隨機分配到 NMN 組(250 mg/天;Oriental Yeast Co., Ltd.,Tokyo,Japan)(圖 S1)。基線測試包括: i) 雙能 X 射線吸收測定法和磁共振成像以評估身體成分; ii) 高胰島素-正常血糖鉗夾程序(4 小時基礎階段和 4 小時 50 mU/m2 /min 的胰島素輸注),結合穩定同位素葡萄糖和棕櫚酸鹽示踪劑輸注,以評估多器官胰島素敏感性; iii) 基礎期股四頭肌經皮活檢和胰島素輸注,以評估 NMN 治療的細胞效應。基線研究完成後,參與者被隨機分配到安慰劑或 NMN 治療 10 週。在參與者完成約 10 週的治療後,重複在基線進行的所有研究程序。此外,在安慰劑或 NMN 攝入後 4 小時評估外周血單個核細胞 (PBMC) NAD+ 含量。根據每週的藥片計數,安慰劑組 99.6% 的處方藥和 NMN 組 100% 的處方藥被參與者服用。兩組均未報告不良事件,也未檢測到標準血液檢查異常(表 S1)。
N-甲基-2-吡啶酮-5-甲酰胺和N-甲基-4-吡啶酮-5-甲酰胺的血漿濃度是NMN產生的代謝物(13),在NMN治療10週後增加,但安慰劑沒有增加(圖1) . 1A)。 NMN 治療 10 週後,PBMC 中的基礎 NAD+ 含量增加,但安慰劑治療後沒有變化(圖 1B)。在治療 10 週結束時攝入單次 250 mg 劑量的 NMN 不會導致 PBMC NAD+ 含量進一步增加至高於基礎水平,這是在攝入後 240 分鐘獲得的連續血液樣本中評估的(圖 1C);然而,由於 NMN 組的基礎值較高,NMN 組的 240 分鐘 PBMC NAD+ 含量高於零的曲線下面積比安慰劑組高 43%(p<0.01)(圖 1D) )。在最後一劑安慰劑或 NMN 後 1.5 小時獲取股四頭肌組織樣本,以評估治療對骨骼肌 NMN 代謝物的影響。儘管兩組的肌肉 NAD+ 和煙酰胺含量在治療 10 週後都沒有變化(圖 1、 、EE 和 和 F)、F),但肌肉 N-甲基-煙酰胺、甲基-2-吡啶酮-5-甲酰胺、和 N-methyl-4-pyridone-5-carboxamide 在 NMN 而不是安慰劑治療後增加(圖 1, ,G,G, ,H,H 和 I),I),表明 NMN 治療增加了肌肉 NAD+周轉。

|
圖1。
血漿、PBMC 和骨骼肌中的 NMN 代謝物和 NAD+。 A,血漿 N-甲基-2-吡啶酮-5-甲酰胺 (2PY) 和 N-甲基-4-吡啶酮-5-甲酰胺 (4PY) 在(白色條)和之後(灰色條)處理之前。 B,基礎外周血單個核細胞 (PBMC) NAD+ 含量在(白條)和處理後(灰條)。 C,在用安慰劑或 NMN 治療 10 週結束時(時間 0)和攝入安慰劑膠囊(白條)或 NMN(250mg)(黑條)後 240 分鐘的 PBMC NAD+ 含量。 D,攝入安慰劑膠囊或 250 mg NMN 後 240 分鐘的 PBMC NAD+ 曲線下面積 (AUC)。 E 到 I,骨骼肌 NAD+、煙酰胺、N-甲基煙酰胺、2PY 和 4PY 含量(白條)和安慰劑或 NMN 治療後(灰色條)。以時間(治療前與治療後)和組(安慰劑與 NMN)為因素的雙向混合模型方差分析 (ANOVA) 用於比較 NMN 和安慰劑治療對基礎 PBMC 和組織 NAD+ 的影響。 Tukey's post-hoc test 之後是一個顯著的組交互時間,以找出顯著的平均差異。使用獨立樣本(雙尾)的 t 檢驗來確定兩組中平均 PBMC AUC 值之間的差異。 *值與相應的治療前值有顯著差異,P <0.01。 **值與相應的治療前值顯著不同,P <0.05。 †值與安慰劑組的相應值顯著不同,P <0.01。圓圈代表個體參與者值:在 11 名安慰劑和 12 名 NMN 參與者中測量了骨骼肌 NAD+ 含量;在 11 名安慰劑和 10 名 NMN 參與者中測量了骨骼肌 NMN 代謝物;所有其他測量均在 12 名安慰劑和 13 名 NMN 參與者中進行。條形代表平均值±SEM。 |
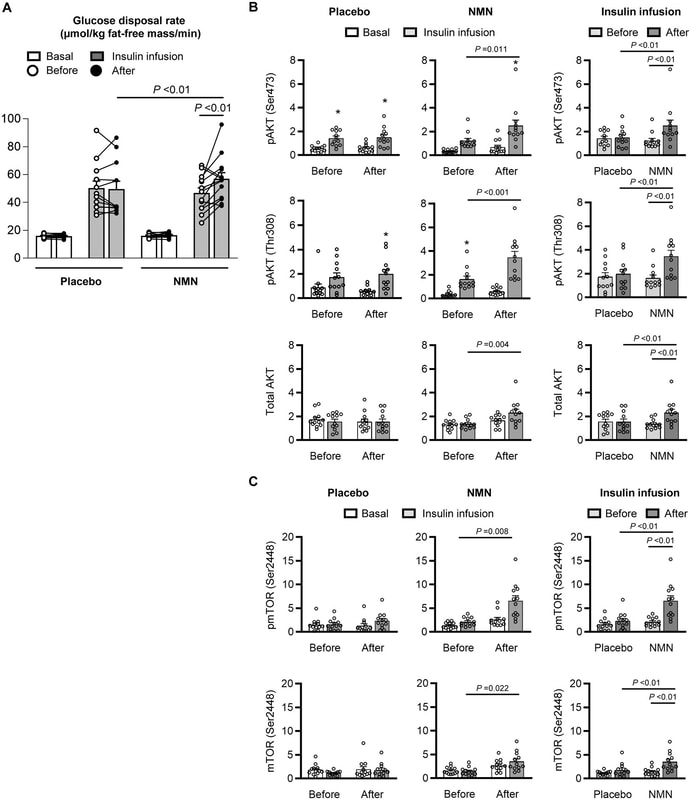
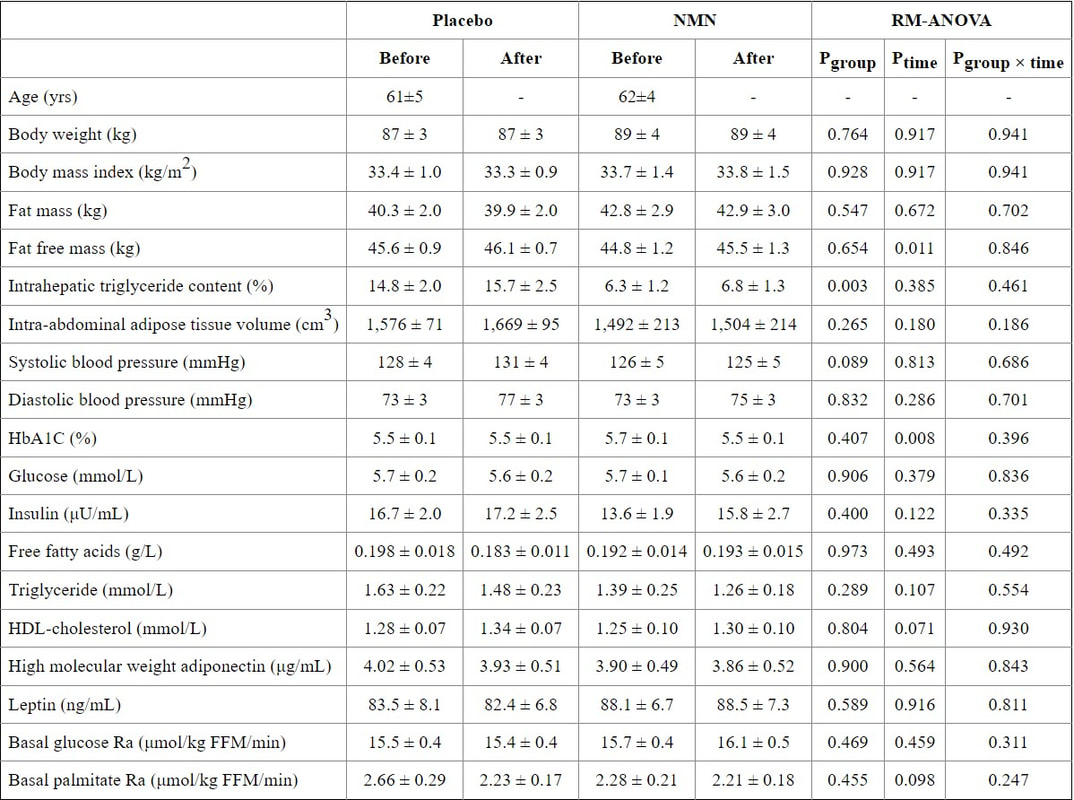
身體成分(脂肪量、無脂肪量、腹內脂肪組織體積和肝內甘油三酯含量)、血壓、血漿葡萄糖、胰島素、游離脂肪酸、脂質、脂聯素和瘦素濃度,以及基礎葡萄糖和脂肪酸治療 10 週後,兩組的動力學均未發生變化(表 1)。肌肉胰島素敏感性,評估為鉗夾手術期間每公斤無脂肪體重的胰島素刺激葡萄糖處理率,在 NMN 補充 10 週後比之前增加 25±7%(p<0.01),但沒有差異安慰劑治療後比安慰劑治療前(圖 2A)。相比之下,肝臟胰島素敏感性(評估為肝臟胰島素敏感性指數(14)和胰島素輸注期間葡萄糖出現率(Ra)的抑制)和脂肪組織胰島素敏感性(評估為脂肪組織胰島素抵抗指數(15)和與安慰劑或 NMN 治療前相比,棕櫚酸 Ra) 的抑制沒有不同(圖 S2)。
|
圖 2。
NMN對骨骼肌胰島素敏感性和信號傳導的影響。 A,肌肉胰島素敏感性,評估為高胰島素-正常血糖鉗夾手術的基礎條件(白條)和胰島素輸注(灰條)期間的葡萄糖處理率。以時間(治療前與治療後)、狀況(基礎與胰島素輸注)和組(安慰劑與 NMN)為因素的三因素混合模型方差分析用於比較 NMN(n=13)和安慰劑(n = 12)治療胰島素刺激的葡萄糖處理率。顯著的三向交互作用(P = 0.022)之後是 Tukey 的事後檢驗以找出顯著的平均差異。平均值的顯著差異由具有相應 P 值的條上方的線表示。 B,在基礎條件和胰島素期間,磷酸化 AKT 絲氨酸 473 (pAKT Ser473)、蘇氨酸 308 (pAKT Thr308) 和 mTOR 絲氨酸 2448 (pmTOR Ser2448) 以及總 AKT 和 mTOR 相對於骨骼肌中的肌動蛋白的蛋白質印跡密度分析和胰島素在安慰劑(n=12)或 NMN(n=12)治療 10 週前後輸注。補充材料中提供了用於單個蛋白質印跡分析的原始圖像(圖 S3)。進行單向方差分析以確定每組內每種時間和條件組合(在每次治療之前和之後基礎和胰島素輸注期間)之間磷酸化 AKT 數量的差異。在一項重要的綜合測試之後,使用 Tukey 的事後測試來定位顯著的平均差異。 C,在胰島素輸注之前(淺灰色條)和之後(深灰色條)安慰劑(n = 12)或NMN (n=12) 治療。以時間(治療前與治療後)和組(安慰劑與 NMN)為因素的雙向混合模型方差分析用於確定變量之間差異的顯著性。顯著的雙向交互之後是 Tukey 的事後檢驗以找出顯著的平均差異。平均值的顯著差異由具有相應 P 值的條上方的線表示。 *值與相應的基礎值顯著不同,P <0.05。圓圈代表個體參與者的價值。條形代表平均值±SEM。 |
|
表一
身體成分和基礎代謝變量
身體成分和基礎代謝變量
值是平均值±SEM。 在安慰劑 (n=12) 或 NMN (n=13) 治療之前和之後測量代謝變量。 使用雙向混合模型方差分析來評估補充 NMN 對每個結果的影響。 NMN 對這些變量的影響與安慰劑沒有什麼不同。 縮寫:FFM,去脂體重
我們評估了補充 NMN 對蛋白激酶 AKT 的絲氨酸 473 (Ser473) 和蘇氨酸 308 (Thr308) 的磷酸化以及肌肉中 mTOR 的 Ser2448 磷酸化的影響,這些是參與調節葡萄糖攝取和調節的胰島素信號通路的關鍵組成部分。肌肉重塑。在 NMN 組中,胰島素輸注期間肌肉 AKT 和 mTOR 磷酸化以及總 AKT 和 mTOR 蛋白豐度在治療後高於治療前,但在安慰劑組中沒有變化(圖 2、B 和 CC 和圖 S3)。這些細胞發現與安慰劑和 NMN 治療對體內觀察到的肌肉胰島素敏感性的影響一致。
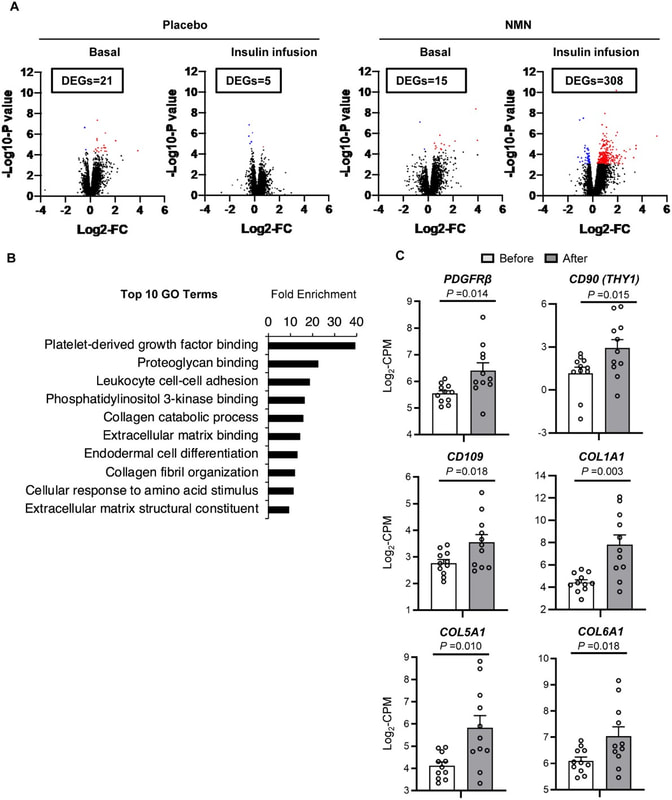
我們使用 RNA 測序來評估股四頭肌樣本的整體基因表達。我們在基礎條件和胰島素輸注期間,在安慰劑或 NMN 治療 10 週之後與之前相比,確定了差異表達的基因(DEG,錯誤發現率 <0.05)(圖 3A 和表 S2)。在基礎條件下,與治療前相比,安慰劑組有 21 個 DEG,NMN 組有 15 個 DEG(圖 3A 和表 S2)。胰島素輸注過程中,安慰劑組和NMN組相比治療前分別增加了5個DEGs和308個DEGs; NMN 組中沒有一個 DEG 受到安慰劑治療的影響(圖 3A 和表 S2)。
我們使用 RNA 測序來評估股四頭肌樣本的整體基因表達。我們在基礎條件和胰島素輸注期間,在安慰劑或 NMN 治療 10 週之後與之前相比,確定了差異表達的基因(DEG,錯誤發現率 <0.05)(圖 3A 和表 S2)。在基礎條件下,與治療前相比,安慰劑組有 21 個 DEG,NMN 組有 15 個 DEG(圖 3A 和表 S2)。胰島素輸注過程中,安慰劑組和NMN組相比治療前分別增加了5個DEGs和308個DEGs; NMN 組中沒有一個 DEG 受到安慰劑治療的影響(圖 3A 和表 S2)。
|
圖 3。
NMN對骨骼肌整體轉錄組譜的影響。 通過使用 RNA 測序 (RNA-seq) 評估在安慰劑 (n=12) 和 NMN 組 (n=11) 治療前後在基礎條件和胰島素輸注期間獲得的骨骼肌組織。 A,骨骼肌 RNA-seq 數據的火山圖,具有 log2 倍變化 (FC)(X 軸)和 -log10-P 值(Y 軸)。在基礎條件和胰島素輸注期間治療前後之間差異表達基因的數量(DEGs,FDR<0.05)顯示在每個面板的方框中。安慰劑組和 NMN 組治療前後顯著上調和下調的 DEG 分別顯示為紅點和藍點。 B,按倍數富集排列的前 10 個基因本體 (GO) 術語。 C,每個參與者在 NMN 治療前後胰島素輸注期間參與 PDGF 信號傳導和肌肉重塑的選擇性蛋白的骨骼肌基因表達。 PDGFRβ、CD90、CD109、COL1A1、COL5A1 和 COL6A1 被鑑定為 DEG(表 S2)。基因表達以每百萬 (CPM) 讀數的 log2 轉換計數表示。線代表 NMN 治療前後個體參與者的值。對配對樣本(雙尾)進行 t 檢驗以確定每個基因在 NMN 處理前後的差異;相應的 P 值顯示在黑線上方。 |
用於註釋、可視化和集成發現途徑分析的數據庫用於識別在 NMN 治療後胰島素輸注期間顯著富集的基因本體 (GO) 術語 (調整後的 P 值 < 0.05)。 “血小板衍生生長因子(PDGF)結合”途徑是最富集的(圖3B和表S3)。 DEGs 在膠原蛋白和細胞外基質代謝的生物學途徑中顯著富集,它們是 PDGF 信號傳導的下游元件(圖 3B 和表 S3)。此外,在胰島素輸注期間,NMN 補充劑顯著上調了 PDGF 受體 β (PDGFRβ) 和骨骼肌週細胞標誌物 (CD90 和 CD109) 的骨骼肌表達(圖 3C)。補充 NMN 還增加了與肌源性 PW1 陽性間質細胞和周細胞(PW1/PEG3、QPRT、NG2 和 PECAM1)相關的其他基因的表達(16、17),但這些增加沒有達到統計學意義(圖 S4 )。此外,PDGF 信號傳導的下游靶標,如 COL1A1、COL5A1 和 COL6A1,在 NMN 處理後顯著上調(圖 3C)。這些結果表明,NMN 治療增加了骨骼肌中的肌原性細胞群並增強了 PDGF 信號傳導。
由於 NAD+ 生物學對線粒體功能的重要性以及肌肉線粒體與肌肉功能之間的關係 (3, 18),我們評估了 NMN 是否影響骨骼肌線粒體呼吸能力和身體功能。通過使用經皮活檢獲得的股四頭肌樣本的高分辨率呼吸測量法評估肌肉線粒體氧化能力,在安慰劑或 NMN 治療 10 週後沒有改變(圖 S5)。通過測量握力和扭矩、疲勞性和從慣用腿的膝伸和屈肌的疲勞運動中恢復來評估的肌肉身體功能不受安慰劑或 NMN 治療 10 週的影響(圖 S6)。
這項隨機、安慰劑對照、雙盲試驗表明,補充 10 週的 NMN 可增加肌肉胰島素信號傳導(增加胰島素刺激的磷酸化 AKT 和 mTOR)和肌肉胰島素敏感性(增加每公斤無脂肪的胰島素刺激的葡萄糖處理率)質量)在絕經後超重或肥胖的前驅糖尿病婦女中。這種肌肉胰島素敏感性的改善具有臨床意義,與肥胖患者體重減輕約 10% (19) 和使用胰島素增敏劑曲格列酮 (20) 治療 12 週後觀察到的改善相似。在我們的參與者中觀察到 NMN 治療後肌肉胰島素刺激的 AKT 磷酸化和葡萄糖攝取的增加與在囓齒動物模型中進行的研究一致 (1, 3, 11)。我們的數據表明,NMN 誘導的肌肉 PDGFRβ 表達增加可能參與介導這種效應,因為 PDGF 信號傳導,特別是通過 PDGFRβ,增強了骨骼肌和多種細胞類型中胰島素刺激的 AKT 磷酸化和葡萄糖轉運 (21-23)。在我們接受 NMN 治療的參與者中,肌肉 NAD+ 的穩態水平沒有變化也與在小鼠中進行的研究一致,這些研究發現 NMN 給藥改善了胰島素敏感性而不改變肌肉 NAD+ 濃度 (11)。然而,NMN 治療增加了肌肉 NMN 代謝物(N-甲基煙酰胺、N-甲基-2-吡啶酮-5-甲酰胺和 N-甲基-4-吡啶酮-5-甲酰胺)的水平,表明 NMN 治療增加了骨骼肌 NAD+周轉。 NMN 對肌肉胰島素敏感性的影響是特異性的,不影響與胰島素抵抗相關的其他重要變量,包括肝臟和脂肪組織胰島素敏感性指數、腹內脂肪組織體積、肝內甘油三酯含量和空腹血糖、胰島素和脂聯素濃度。這些結果表明,NMN 對骨骼肌中胰島素介導的葡萄糖代謝具有選擇性的有益作用。
我們的數據證明了 NMN 對骨骼肌生物學的強大影響。從我們對骨骼肌的無偏全球轉錄組分析獲得的數據表明,與安慰劑相比,NMN 在胰島素輸注過程中導致 DEG 數量增加了 60 倍。 PDGF 結合途徑富含 DEG,肌肉 PDGFRβ 和 PDGF 信號通路的下游靶點在 NMN 治療後的胰島素輸注過程中顯著上調。此外,NMN 治療增加了胰島素刺激的 mTOR 磷酸化和肌源性 PW1/PEG3 陽性間質細胞和周細胞的選定標誌物的基因表達 (16, 17),這共同表明骨骼肌重塑和再生增加。
與我們在參與者中觀察到的 NMN 的胰島素增敏作用相比,在中年和老年男性中進行的幾項隨機對照試驗的結果發現,使用煙酰胺核苷 (NR) 進行治療,這是另一種可提高胰島素敏感性的 NAD+ 中間體在囓齒動物 (5, 8, 24) 中,不影響全身或肌肉胰島素敏感性 (25-27)。在這些臨床研究中補充 NR 缺乏代謝益處的原因尚不清楚,需要進一步的研究來解決這個問題。
我們的研究結果表明,補充 NMN(250 毫克/天)可增加超重或肥胖的絕經後前驅糖尿病婦女的骨骼肌胰島素信號傳導、胰島素敏感性和肌肉重塑。導致這些代謝效應的確切機制以及補充 NMN 在其他患者群體中的潛在代謝益處仍有待探索。
由於 NAD+ 生物學對線粒體功能的重要性以及肌肉線粒體與肌肉功能之間的關係 (3, 18),我們評估了 NMN 是否影響骨骼肌線粒體呼吸能力和身體功能。通過使用經皮活檢獲得的股四頭肌樣本的高分辨率呼吸測量法評估肌肉線粒體氧化能力,在安慰劑或 NMN 治療 10 週後沒有改變(圖 S5)。通過測量握力和扭矩、疲勞性和從慣用腿的膝伸和屈肌的疲勞運動中恢復來評估的肌肉身體功能不受安慰劑或 NMN 治療 10 週的影響(圖 S6)。
這項隨機、安慰劑對照、雙盲試驗表明,補充 10 週的 NMN 可增加肌肉胰島素信號傳導(增加胰島素刺激的磷酸化 AKT 和 mTOR)和肌肉胰島素敏感性(增加每公斤無脂肪的胰島素刺激的葡萄糖處理率)質量)在絕經後超重或肥胖的前驅糖尿病婦女中。這種肌肉胰島素敏感性的改善具有臨床意義,與肥胖患者體重減輕約 10% (19) 和使用胰島素增敏劑曲格列酮 (20) 治療 12 週後觀察到的改善相似。在我們的參與者中觀察到 NMN 治療後肌肉胰島素刺激的 AKT 磷酸化和葡萄糖攝取的增加與在囓齒動物模型中進行的研究一致 (1, 3, 11)。我們的數據表明,NMN 誘導的肌肉 PDGFRβ 表達增加可能參與介導這種效應,因為 PDGF 信號傳導,特別是通過 PDGFRβ,增強了骨骼肌和多種細胞類型中胰島素刺激的 AKT 磷酸化和葡萄糖轉運 (21-23)。在我們接受 NMN 治療的參與者中,肌肉 NAD+ 的穩態水平沒有變化也與在小鼠中進行的研究一致,這些研究發現 NMN 給藥改善了胰島素敏感性而不改變肌肉 NAD+ 濃度 (11)。然而,NMN 治療增加了肌肉 NMN 代謝物(N-甲基煙酰胺、N-甲基-2-吡啶酮-5-甲酰胺和 N-甲基-4-吡啶酮-5-甲酰胺)的水平,表明 NMN 治療增加了骨骼肌 NAD+周轉。 NMN 對肌肉胰島素敏感性的影響是特異性的,不影響與胰島素抵抗相關的其他重要變量,包括肝臟和脂肪組織胰島素敏感性指數、腹內脂肪組織體積、肝內甘油三酯含量和空腹血糖、胰島素和脂聯素濃度。這些結果表明,NMN 對骨骼肌中胰島素介導的葡萄糖代謝具有選擇性的有益作用。
我們的數據證明了 NMN 對骨骼肌生物學的強大影響。從我們對骨骼肌的無偏全球轉錄組分析獲得的數據表明,與安慰劑相比,NMN 在胰島素輸注過程中導致 DEG 數量增加了 60 倍。 PDGF 結合途徑富含 DEG,肌肉 PDGFRβ 和 PDGF 信號通路的下游靶點在 NMN 治療後的胰島素輸注過程中顯著上調。此外,NMN 治療增加了胰島素刺激的 mTOR 磷酸化和肌源性 PW1/PEG3 陽性間質細胞和周細胞的選定標誌物的基因表達 (16, 17),這共同表明骨骼肌重塑和再生增加。
與我們在參與者中觀察到的 NMN 的胰島素增敏作用相比,在中年和老年男性中進行的幾項隨機對照試驗的結果發現,使用煙酰胺核苷 (NR) 進行治療,這是另一種可提高胰島素敏感性的 NAD+ 中間體在囓齒動物 (5, 8, 24) 中,不影響全身或肌肉胰島素敏感性 (25-27)。在這些臨床研究中補充 NR 缺乏代謝益處的原因尚不清楚,需要進一步的研究來解決這個問題。
我們的研究結果表明,補充 NMN(250 毫克/天)可增加超重或肥胖的絕經後前驅糖尿病婦女的骨骼肌胰島素信號傳導、胰島素敏感性和肌肉重塑。導致這些代謝效應的確切機制以及補充 NMN 在其他患者群體中的潛在代謝益處仍有待探索。
